

Sensing touch movement and sound
Part 4: Scolopidia and chordotonal organs
The third type of mechanoreceptor is the scolopidium. This is sometimes referred to as a chordotonal organ. In its simplest form it is a single bipolar neuron with specialized structure that connects to the epidermis beneath the cuticle. In some insects the scolopidia are clustered together into complex hearing organs. Others such as moths have two neurons that connect with the ear beneath each wing. Scolopidia are used for hearing sound, sensing movement, pressure and position.
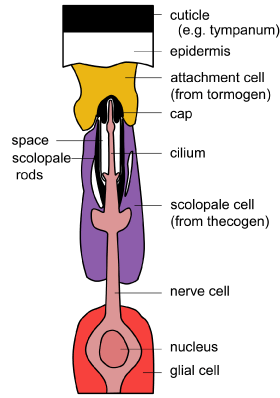 |
There are three components to the basic structure of a scolopidium: a neuron, an enveloping (scolopale) cell and an attachment cell. Scolopidia are attached to the epidermis of the cuticle at one or both ends. Some scolopidia contain more than one neuron. The term chordotonal organ is sometimes used instead of scolopidium or scolopidia. [Diagram: BW Cribb. Modified from Chapman edition 4] |
The sensory neuron extends as a narrow cilium-like process akin to the dendrite in other mechanoreceptors. However, this cilium process contains a ring structure of nine double filaments and a proximal ciliary root. The tip of the process is encased in a cap and this in turn is embedded in the attachment cell that connects with the epidermis of the cuticle. Below the cap is a complex scolopale cell that contains rods and an extracellular space. The scolopale cell surrounds the cilium-like process. Beneath this sits a sheath cell. A glial cell lies proximally around the axon.
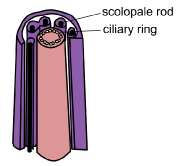 |
Schematic diagram of cilium structure with the ciliary ring and the surrounding scolopale rods. [Diagram BW Cribb] |
The displacement of the cap and sliding of the tubule doublets within the cilium are implicated in stimulation of the neuron. Stretch-activation is involved (see section on mechanism). Some scolopidia run between segments of the cuticle and are connected at both ends. These inform the insect about body movement.
Scolopidia, either singly or in groups, are responsible for monitoring head to body positions, monitoring movement of adjacent body segments, sensing gut expansion, wing forces, reproductive system pressures, vibration of antennae, and hearing pressure changes in the air (sound).
Chordotonal organs
Subgenual organ
This is an internal organ made up of 10-40 Scolopidia and present in the proximal part of the tibia. It is absent in Coleoptera and Diptera. The cluster of sensory neurons attach to the cuticle and respond to substrate vibrations, for example at low frequencies up to 50Hz and at high frequencies up to 8kHz depending on the species investigated. These signals are important for detection of predators and also intraspecific signals (that is from mates or rivals).

[Diagram: BW Cribb. Modified from Chapman edition 4: the subgenual organ of an ant]
Johnston’s organ
The Johnston’s organ is positioned at the base of the antennae. It occurs in the pedicel and monitors vibration of the antennal flagellum.
 |
Head of a biting midge: Forcipomyia (Lasiohelea) townsvillensis showing the antennae and basal pedicel containing the Johnston’s organ [Image: BW Cribb] |
The scolopidia are present either as a single mass or in several groups. The organ is best known from the mosquito ear used by male mosquitoes to hear the wing beat frequency of females. Here, the scolopidia are all connected in a ring to a basal plate positioned around the end of the antennal flagellum. But it occurs in all adult insects and in many larvae. In grasshoppers, bees, butterflies and some flies it acts as an air speed monitor. In Drosophila it monitors near-field sound.
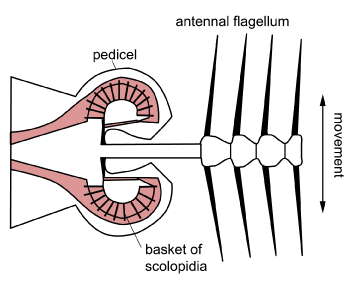
[Diagram: BW Cribb]
Tympanal organs
True ears in insects are composed of a thin cuticular membrane (disc or tympanic membrane) backed by an air sac so it can vibrate freely with changes in air pressure. From one to over 1000 (in cicadas) scolopidia are attached to the centre or adjacent to the tympanic membrane. Tympanal organs occur in a variety of positions. In crickets they occur on the legs (fore tibia).
|
Cricket’s leg: scanning electron micrograph [Image: BW Cribb] |
Close up of ear on cricket’s leg [Image BW Cribb] |
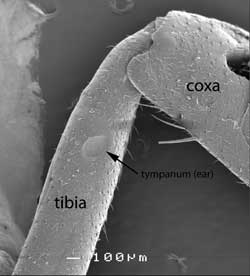
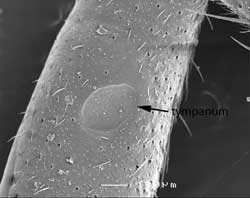
In grasshoppers and cicadas the “ears” are found in the first abdominal segment.
 |
In noctuid moths they occur in the posterior part of the metathorax under the wings as shown in this image of Helicoverpa armigera. [Image BW Cribb] |
In Chrysopa they occur in the wing (ventral side of the radial vein of the fore wing) and in this case the tympanum is not backed by an air space but rather by hemolymph.
Tympanal organs work by transducing sound pressure. The membrane is displaced back and forth with sound. The vibrations vary with the intensity of the sound and the structure of the membrane. The sensory neuron is stimulated by the membrane displacement. Number of action potentials generated is generally proportional to the loudness: the louder the sound, the more action potentials are sent.
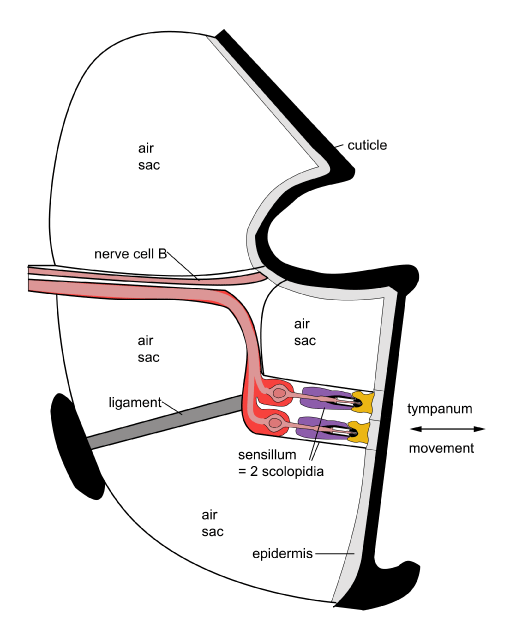
In noctuid moths there are two scolopidia and each responds to sound in a different way which allows the monitoring of distant and close sounds and allows effective escape from bat predators.
The two tympanal nerves (attached to the scolopidia) are termed A1 and A2. A1 requires bursts of sound to keep it activated. These are found in the sonic calls of bats. Other sounds that are longer-lasting result in the nerve shutting down response since it responds in a phasic manner. As the sound becomes louder, that is, the bat gets closer, A1 fires more frequently. Only when the sound is very loud does the A2 cell start firing. When this second cell fires the moth flight turns irregular and it dives, enabling it to escape a bat encounter.
Sound is sent between insects too. In this instance it is often dependant on pulsing. Cicadas and crickets pulse sound and can separate pulses up to about 100Hz. Here we see communication based on sound patterns rather than frequency. Direction is aided by pairing of tympanal organs where they occur on different sides of the body