

Sensing Touch, movement and sound
Part 2: Setae
The most visible sense organs involved in mechanoreception (touch) are stout hairs that taper to a fine tip. The principle feature is a socket in which the hair shaft sits. As the hair bends in this socket it stimulates a specialized nerve ending at the base. The nerve cell is bipolar. The correct term for these bristles is sensillum chaeticum (singular) or sensilla chaetica (plural). Finer hairs are called sensilla trichodea (plural), which means sensory hair.
These touch-sensitive hairs are the most visible feature of insects. They are found on all body parts: legs, wings, ovipositors, head and its appendages, general body segments and even on the eyes of some insects. The tarsi of insects always have mechanoreceptive setae present. In this scanning electron microscope image (right) we see a tree-like pulvillus in the centre of two curved claws. These are not mechanoreceptors. But the surrounding stout spines are touch-sensitive. [Image BW Cribb] |
 |
|
Left: Caterpillar (Helicoverpa armigera) with touch-sensitive hairs across the body [Image BW Cribb] |
Spines on the head and wings also enable a winged insect to sense the rush of wind, helping it to monitor when it is flying. Right: At low magnification, under a scanning electron microscope, we can see the setae arranged on the wing veins of a lacewing. [Image BW Cribb] |
 |
 |
Sometimes the hair can bend in any direction, as in the bristles on wings. This image (left) shows a mechanoreceptive seta on the wing of a lacewing. Note the socket. [Image BW Cribb] |

The sensillum contains a bipolar neuron at the base. It is positioned at one side of the socket and is stimulated by the bending and stretching that occurs when the hair shaft is moved. The nerve cell projects upwards to form a dendrite and ends in what is termed a ‘tubular body’. This body is particularly characteristic of insect mechanoreceptors associated with cuticular structures like hairs and domes. It is so called because when observed using transmission electron microscopy, the ending is packed full of tubules arranged parallel to one another. A sheath surrounds the dendrite (termed the dendrite sheath) and also the tubular body and connects into the base of the hair shaft. The innermost sheath cell (thecogen) secretes the dendrite sheath. The outermost sheath cell (tormogen) secretes the socket before withdrawing to leave a lymph chamber behind.
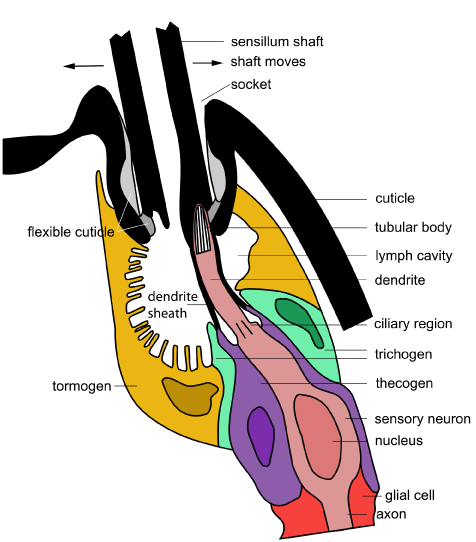
This diagram of a mechanoreceptive seta is based on those found in biting midges but the form is generic across all insects [Diagram: BW Cribb]
The neuron is surrounded by accessory cells. These are the inner thecogen, the middle trichogen and the outer tormogen. They play a similar role to the accessory cells in chemoreceptors. In the diagram here, the cells have been colour-coded so you can follow them across the different diagrams in this section and compare them with the diagrams in the section on chemoreceptors.
Directional sensitivity is the result of the position where the dendrite joins the shaft. Movement of the shaft in the socket transfers as stress on the tubular body. This results in the generation of a receptor potential. At a certain threshold, a series of nerve impulses (action potentials) are sent from the sensory neuron down the axon to the central nervous system.
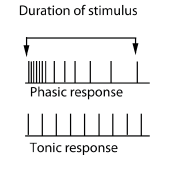 |
Impulses may be sent only with the initial bending of the hair. With continued deflection of the hair the nerve signals stop. This is known as a ‘phasic response’. The other type of response is a ‘tonic response’. Here the action potentials continue to be sent as long as the deflection of the hair continues. Some receptors have both a phasic and a tonic component to their response. |
Hairs responsible for sensing touch usually respond in a phasic manner (starting with action potentials closely spaced in time and then these become less and less frequent). This is because they inform the insect about ‘new’ interactions.
Beds of hairs that monitor body position (proprioception) respond with a mixed phasic/tonic response.
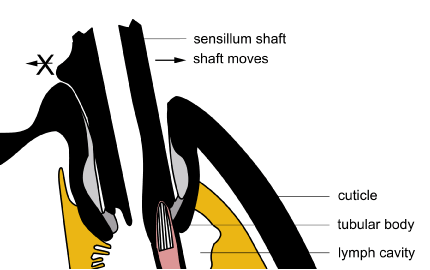
Whereas most bristles can move in any direction, some can only bend in one direction because they are locked from moving back by a protrusion of the shaft and the way the socket is designed. An example is the raster in cane beetle grubs – which is a collection of setae near the anus (see Cribb et al. 1998. Structure of raster in melolonthine larvae. Annals of the Entomological Society of America 91(2), 202-210).
In this diagram, the shaft has been modified to stop its movement to the left [Diagram BW Cribb].
The spines that enable sensing of touch are spread across the body. Some are also clustered together into fields at the joints of insects and allow the sensing of leg, segment, or head movement. As the body components move against one another they rub or press against these hair plates (fields of hairs). Monitoring body position is termed proprioception.
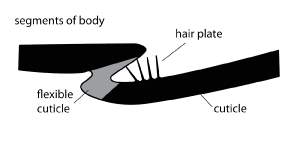
[Diagram BW Cribb]
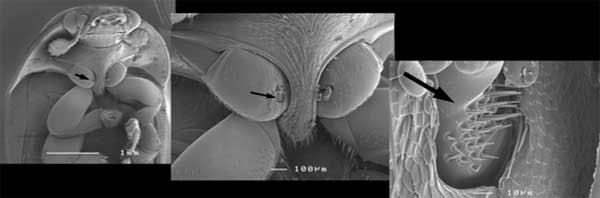
This is the ventral surface of a small hive beetle, Athena tumida (Coleoptera: Nitidulidae), a pest of beehives in Australia, photographed using a scanning electron microscope. The arrow indicates the position of a hair plate at the base of a coxa. When the beetle walks the hair plate will rub against the sternum nearby and act as a proprioceptor. The small balls on the insect surface are corn starch, through which the insect has been crawling. [Photo B. Cribb]